Pergerakan haiwan
Pergerakan (lokomotor) haiwan dalam etologi merujuk kepada mana-mana kaedah yang digunakan haiwan untuk bergerak dari satu tempat ke tempat lain.[1] Beberapa mod pergerakan ialah (pada mulanya) gerakan sendiri, termasuk berlari, berenang, melompat, terbang, melompat, melambung dan meluncur. Terdapat juga banyak spesies haiwan yang bergantung pada persekitaran mereka untuk bergerak, sejenis mobiliti yang dipanggil sebagai pergerakan pasif, contohnya, belayar (sesetengah obor-obor), berguling (beberapa kumbang dan labah-labah) atau menunggang haiwan lain.

Haiwan bergerak atas pelbagai sebab, seperti mencari makanan, pasangan, habitat mikro yang sesuai atau untuk melarikan diri daripada pemangsa. Bagi kebanyakan haiwan, keupayaan untuk bergerak adalah penting untuk kelangsungan hidup dan, akibatnya, pemilihan semula jadi telah membentuk kaedah dan gaya pergerakan yang digunakan oleh organisma. Contohnya, haiwan migrasi yang menempuh jarak yang jauh lazimnya mempunyai gaya pergerakan yang menelan kos tenaga yang sangat sedikit bagi setiap unit jarak, manakala haiwan bukan migrasi yang mesti kerap bergerak pantas untuk melarikan diri daripada pemangsa memiliki gaya gerakan yang mungkin menelan tenaga yang tinggi, tetapi sangat pantas.
Struktur anatomi yang digunakan haiwan untuk pergerakan, termasuk silia, kaki, sayap, lengan, sirip atau ekor kadangkala dirujuk sebagai organ lokomotor[2] atau struktur lokomotor.[3]
Pergerakan dalam media berbeza sunting
Haiwan bergerak melalui, atau melalui, empat jenis persekitaran: akuatik (dalam atau di atas air), daratan (di atas tanah atau permukaan lain, termasuk arboreal, atau kediaman pokok), fossorial (bawah tanah) dan udara. Banyak haiwan—contohnya haiwan separa akuatik dan burung penyelam— selalu bergerak melalui lebih daripada satu jenis medium. Dalam sesetengah kes, permukaan yang mereka gerakkan memudahkan kaedah pergerakan mereka.
Akuatik sunting
Berenang sunting

Di kawasan air, mengapungkan diri adalah mungkin menggunakan daya apungan. Jika badan haiwan kurang tumpat daripada air, ia boleh kekal terapung. Ini memerlukan sedikit tenaga untuk mengekalkan kedudukan menegak, tetapi memerlukan lebih banyak tenaga bagi pergerakan satah mendatar berbanding dengan haiwan yang kurang apung. Daya seretan yang dialami di dalam air adalah lebih besar daripada udara. Oleh itu, morfologi adalah penting bagi pergerakan yang cekap, dan dalam kebanyakan kes, penting bagi fungsi asas seperti menangkap mangsa. Bentuk badan fusiform, seperti torpedo dilihat pada kebanyakan haiwan akuatik,[4][5] walaupun mekanisma yang digunakan dalam pergerakan adalah pelbagai.
Cara utama ikan menghasilkan tujahan adalah dengan mengayunkan badan dari sisi ke sisi, dengan gerakan gelombang yang terhasil berakhir pada sirip ekor yang besar. Kawalan yang lebih halus seperti untuk pergerakan perlahan, selalunya dicapai dengan tujahan dari sirip dada (atau anggota hadapan mamalia laut). Sesetengah ikan, sperti ikan Hydrolagus colliei dan ikan batiform (ikan gergaji, ikan gitar dan pari) menggunakan sirip dada mereka sebagai alat pergerakan utama, kadangkala dipanggil sebagai renang labriform. Mamalia marin mengayunkan badannya ke arah atas dan bawah (dorsoventral). Haiwan lain, contohnya penguin dan itik menyelam, bergerak di bawah air dengan cara yang dipanggil sebagai "terbangan akuatik".[6] Sesetengah ikan mendorong diri mereka sendiri tanpa gerakan gelombang badan, seperti kuda laut bergerak perlahan dan Gymnotus.[7]
Haiwan lain seperti sefalopod, menggunakan pendorong jet untuk bergerak pantas, dengan mengambil air dan memancutkannya keluar dalam suatu letupan.[8] Haiwan berenang lain mungkin bergantung terutamanya pada anggota badan mereka, sama seperti manusia semasa berenang. Walaupun hidupan di darat berasal dari laut, haiwan darat telah kembali kepada gaya hidup akuatik pada beberapa kali, seperti setasea akuatik penuh, yang kini, sangat berbeza daripada leluhur darat mereka.
Ikan lumba-lumba kadangkala menunggang ombak haluan yang dicipta oleh bot atau melayari ombak yang memecah secara semula jadi.[9]
Udara sunting
Penerbangan aktif sunting

Graviti ialah halangan utama penerbangan. Oleh kerana mustahil mana-mana organisma mempunyai ketumpatan serendah udara, haiwan terbang mesti menjana daya angkat yang mencukupi untuk naik dan kekal di udara. Satu cara untuk mencapainya ialah dengan sayap, yang digerakkan melalui udara lalu menghasilkan daya angkat ke atas pada badan haiwan. Haiwan yang terbang mestilah sangat ringan untuk mencapai penerbangan, haiwan terbang hidup terbesar ialah burung dengan berat sekitar 20 kilogram.[10] Penyesuaian struktur lain haiwan terbang termasuk berat badan yang dikurangkan dan diagihkan semula, bentuk fusiform dan otot penerbangan yang kuat;[11] dan mungkin juga terdapat penyesuaian fisiologi.[12] Penerbangan aktif telah berevolusi secara bebas sekurang-kurangnya empat kali, dalam serangga, pterosaurus, burung dan kelawar. Serangga ialah takson pertama yang mengembangkan penerbangan, kira-kira 400 juta tahun dahulu (mya),[13] diikuti oleh pterosaurus pada kira-kira 220 mya,[14] burung sekitar 160 mya,[15] dan kelawar pada kira-kira 60 mya.[16]
Terrestrial sunting
Bentuk pergerakan di darat termasuk berjalan, berlari, meloncat atau melompat, menyeret dan merangkak atau merayap. Di sini, geseran dan daya apungan tidak lagi menjadi isu, tetapi struktur rangka dan otot yang kuat diperlukan dalam kebanyakan haiwan darat. Setiap langkah juga memerlukan banyak tenaga untuk mengatasi inersia, dan haiwan boleh menyimpan tenaga potensi anjal dalam tendon mereka untuk membantu mengatasinya. Imbangan juga diperlukan dalam pergerakan darat. Bayi manusia belajar merangkak terlebih dahulu sebelum berupaya berdiri, yang memerlukan koordinasi yang baik serta perkembangan fizikal. Manusia adalah haiwan berkaki dua, berdiri dengan dua kaki serta satu kaki di atas tanah sepanjang masa semasa berjalan. Apabila berlari, hanya satu kaki berada di atas tanah pada satu-satu masa, dan kedua-duanya meninggalkan tanah untuk seketika. Pada kelajuan yang lebih tinggi, momentum membantu mengekalkan badan tegak, jadi lebih banyak tenaga boleh digunakan dalam pergerakan.
Melompat sunting
Melompat boleh dibezakan daripada berlari, dan gaya gerakan lain, di mana seluruh badan dibawa ke udara buat sementara waktu dengan tempoh fasa udara yang agak panjang, dan sudut tinggi pelancaran awal. Banyak haiwan darat melompat untuk melarikan diri daripada pemangsa, atau menangkap mangsa— namun, agak sedikit haiwan menggunakan ini sebagai cara pergerakan utama. Haiwan melompat utama termasuk kanggaru dan makropod lain: arnab, kelinci, jerboa, tikus melompat dan tikus kanggaru. Tikus kanggaru sering melompat 2 m[17] dan dilaporkan mencecah 2.75 m[18] pada kelajuan sehingga hampir 3 m/s (6.7 mph).[19] Mereka boleh menukar arah dengan cepat dalam lompatan.[19] Penggunaan mod "gerak-beku" juga boleh menjadikannya kurang tampak kepada pemangsa malam.[20] Katak adalah, berbanding saiznya, pelompat terbaik dari semua vertebrat.[21] Katak roket Australia, Litoria nasuta, boleh melompat lebih 2 meter (6 ka 7 in), lebih 50 kali panjang badannya.[22]
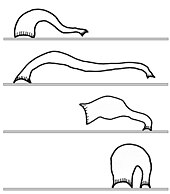
Peristalsis dan bergelung sunting
Haiwan lain bergerak di habitat darat tanpa bantuan kaki. Cacing tanah merangkak dengan peristalsis, suatu kontraksi berirama sama yang mendorong makanan melalui saluran pencernaan manusia.[23]
Berjalan dan berlari sunting
Spesies mempunyai bilangan kaki yang berbeza menyebabkan perbezaan besar dalam pergerakan.
Burung moden, walaupun dikelaskan sebagai tetrapod, biasanya hanya mempunyai dua kaki berfungsi, yang sesetengahnya (cth, burung unta, emu, kiwi) digunakan sebagai mod pergerakan utamanya, yakni bipedalisme. Beberapa spesies mamalia moden adalah biped lazim, iaitu, kaedah pergerakan normalnya ialah jalan berkaki dua. Ini termasuk makropod, tikus serta tikus kanggaru, tikus melompat, pangolin dan beruk hominid. Bipedalisme jarang ditemui di luar haiwan darat— walaupun sekurang-kurangnya dua jenis sotong berjalan secara dua kaki di dasar laut dengan dua lengan mereka, dan berupaya menggunakan lengan-lengan lain untuk menyamar sebagai tikar alga atau kelapa terapung.[24]
Haiwan berkaki tiga tidak wujud— walaupun beberapa makropod seperti kanggaru berselang-seli antara meletakkan berat badan mereka pada ekor berotot serta dua kaki belakang, dan boleh dilihat sebagai contoh pergerakan tripedal dalam haiwan.
Banyak haiwan lazim adalah berkaki empat, dan berjalan atau berlari dengan empat kaki. Beberapa ekor burung menggunakan pergerakan berkaki empat dalam beberapa keadaan. Contohnya, burung kasutkadangkala menggunakan sayapnya untuk menegakkan dirinya selepas menerjang mangsa.[25] Burung hoatzin yang baru menetas mempunyai kuku pada ibu jari dan jari pertama, dan membolehkannya memanjat dahan pokok dengan cekap sehingga sayapnya cukup kuat bagi penerbangan yang berterusan.[26] Kuku ini hilang apabila burung itu mencapai usia dewasa.
Sebilangan kecil haiwan menggunakan lima anggota badan untuk pergerakan. Haiwan berkaki empat boleh menggunakan ekor mereka untuk membantu pergerakan dan semasa merumput; kanggaru dan makropod lain menggunakan ekor mereka untuk mendorong diri ke hadapan dengan empat kaki digunakan untuk mengekalkan keseimbangan.
Serangga biasanya berjalan dengan enam kaki— walaupun sesetengah serangga seperti kupu-kupu nimfalid[27] tidak menggunakan kaki hadapan untuk berjalan.
Araknid mempunyai lapan kaki. Kebanyakan araknid kekurangan otot ekstensor pada sendi distal apendaj. Labah-labah dan ketungging memanjangkan anggota badan mereka secara hidraulik menggunakan tekanan hemolimfa.[28] Solifugae dan beberapa spesies Opiliones memanjangkan lutut mereka dengan menggunakan tebalan elastik dalam kutikula sendi.[28] Kala jengking sebagai contoh, memiliki beberapa otot yang memanjangkan dua sendi kaki (sendi femur-patella dan patella-tibia) serentak.[29][30]
Bawah tanah sunting
Sesetengah haiwan bergerak melalui pepejal seperti tanah dengan menggali menggunakan peristalsis, seperti dalam cacing tanah,[31] atau kaedah lain. Dalam pepejal longgar seperti pasir, sesetengah haiwan dapat bergerak dengan lebih pantas, dengan "berenang" melalui substrat yang longgar. Haiwan menggali termasuk tupai tanah dan sesorok.
Arboreal sunting

Pergerakan arboreal merujuk kepada pergerakan haiwan di pokok. Sesetengah haiwan hanya boleh menaiki pokok sekali-sekala, manakala yang lain adalah arboreal secara eksklusif. Habitat ini menimbulkan pelbagai cabaran mekanikal kepada haiwan yang bergerak melaluinya, dan membawa kepada pelbagai kesan anatomi, tingkah laku dan ekologi serta variasi di seluruh spesies yang berbeza.[32] Tambahan pula, banyak prinsip yang sama ini boleh digunakan untuk memanjat tanpa pokok, seperti di atas timbunan batu atau gunung. Tetrapod yang paling awal diketahui dengan pengkhususan yang menyesuaikannya untuk memanjat pokok ialah Suminia, sinapsid zaman Permian, kira-kira 260 juta tahun dahulu.[33] Sesetengah haiwan invertebrat adalah arboreal eksklusif dalam habitat, contohnya, siput pokok.
Pergerakan pasif sunting
Pergerakan pasif dalam haiwan ialah sejenis mobiliti di mana haiwan itu bergantung pada persekitarannya untuk pengangkutan; haiwan tersebut tidak bergerak tetapi tidak motil.[1]
Hidrozoa sunting
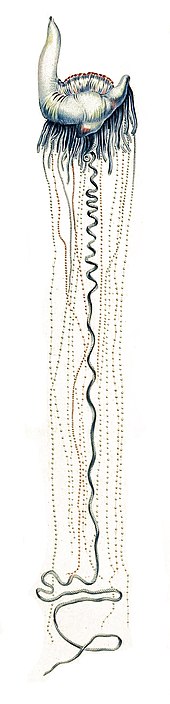
Haiwan man o' war (Physalia physalis) tinggal di permukaan lautan. Pundi dipenuhi gas, atau pneumatofor, kekal di permukaan, manakala selebihnya tenggelam. Oleh kerana man o' war tidak mempunyai alat pendorong, ia digerakkan oleh gabungan angin, arus, dan pasang surut. Pundi pula dilengkapi dengan sifon. Sekiranya berlaku serangan permukaan, pundi iniboleh dikempiskan, membolehkan organisma tenggelam seketika.[34]
Pengangkutan haiwan sunting
Sesetengah haiwan dapat bergerak kerana melekat pada, atau tinggal di atas, haiwan lain atau struktur bergerak. Ini boleh dikatakan lebih tepat dipanggil "pengangkutan haiwan".
Remoras sunting

Ikan remora ialah keluarga (Echeneidae) ikan bersirip sinar. Besarnya sekitar 30–90 cm (0.98–2.95 ka) panjang, dan sirip dorsal pertamanya diubah suai dengan struktur seperti penyedut yang membuka tutup untuk menghasilkan sedutan dan memegang kuat pada kulit haiwan laut yang lebih besar.[35] Dengan menggelongsor ke belakang, remora boleh meningkatkan sedutan, atau ia boleh melepaskan dirinya dengan berenang ke hadapan. Remora kadangkala melekat pada bot kecil. Ikan ini mampu berenang dengan baik secara sendiri, dengan gerakan berliku-liku, atau melengkung. Apabila remora mencapai kira-kira 3 cm (1.2 in), sirip ini terbentuk sepenuhnya dan remora kemudiannya boleh melekat pada haiwan lain. Rahang bawah remora menonjol di luar bahagian atas, dan haiwan itu tidak mempunyai pundi. Sesetengah remora dikaitkan terutamanya dengan spesies hos tertentu. Ia biasanya ditemui melekat pada jerung, pari manta, ikan paus, penyu, dan dugong. Remora yang lebih kecil juga terikat pada ikan seperti tuna dan todak, dan beberapa remora kecil bergerak di dalam mulut atau insang pari manta besar, ikan matahari laut, ikan todak dan ikan layar. Remora mendapat manfaat dengan menggunakan hos sebagai pengangkutan dan perlindungan, dan juga memakan bahan yang dibuang hos.
Parasit sunting
Banyak parasit diangkut oleh perumahnya. Sebagai contoh, endoparasit seperti cacing pita hidup dalam saluran makanan haiwan lain, dan bergantung kepada keupayaan pergerakan perumah untuk menyebarkan telur mereka. Ektoparasit seperti kutu boleh bergerak di atas badan perumahnya, tetapi diangkut dengan jarak yang lebih jauh oleh pergerakan perumah. Sesetengah ektoparasit seperti kutu boleh menumpang dengan cepat (foresis) dan cuba mencari perumah baharu.[36]
Pertukaran antara medium sunting
Sesetengah haiwan bergerak antara medium yang berbeza, cth, dari akuatik ke udara. Ini selalunya memerlukan mod pergerakan yang berbeza dalam persekitaran berbeza, dan mungkin memerlukan gaya gerak peralihan yang khusus.
Terdapat sejumlah besar haiwan separa akuatik (haiwan yang menghabiskan sebahagian daripada kitaran hidup mereka di dalam air, atau secara amnya mempunyai sebahagian daripada anatomi mereka di bawah air). Ini mewakili taksa utama mamalia (cth, memerang, memerang, beruang kutub), burung (cth, penguin, itik), reptilia (cth, anaconda, penyu rawa, iguana marin) dan amfibia (cth, salamander, katak, kadal air).
Rujukan sunting
- ^ a b "Animal locomotion". Encyclopædia Britannica. Dicapai pada December 16, 2014.
- ^ Beckett, B. S. (1986). Biology: A Modern Introduction. Oxford University Press. m/s. 307. ISBN 9780199142606.
- ^ Lindsay, Everett H.; Fahlbusch, Volker; Mein, Pierre (2013), European Neogene Mammal Chronology, Nato Science Series A, 180, Springer Science & Business Media, m/s. 658, ISBN 9781489925138
- ^ Gaston, K.A., Eft, J.A. and Lauer, T.E. (2016). "Morphology and its effect on habitat selection of stream fishes". Proceedings of the Indiana Academy of Science. 121 (1): 71–78.CS1 maint: multiple names: authors list (link)
- ^ Dewar, H.; Graham, J. (1994). "Studies of tropical tuna swimming performance in a large water tunnel-kinematics". Journal of Experimental Biology. 192 (1): 45–59. doi:10.1242/jeb.192.1.45. PMID 9317308.
- ^ Walker, J.A. and Westneat, M.W. (2000). "Mechanical performance of aquatic rowing and flying". Proceedings of the Royal Society of London B: Biological Sciences. 267 (1455): 1875–1881. doi:10.1098/rspb.2000.1224. PMC 1690750. PMID 11052539.CS1 maint: multiple names: authors list (link)
- ^ Sfakiotakis, M., Lane, D.M. and Davies, J.B.C. (1999). "Review of Fish Swimming Modes for Aquatic Locomotion" (PDF). IEEE Journal of Oceanic Engineering. 24 (2): 237–252. Bibcode:1999IJOE...24..237S. CiteSeerX 10.1.1.459.8614. doi:10.1109/48.757275. Diarkibkan daripada yang asal (PDF) pada 2013-12-24.CS1 maint: multiple names: authors list (link)
- ^ Young, R.E.; Katharina M. Mangold, K.M. "Cephalopod jet propulsion". Tree of Life. Dicapai pada October 16, 2016.
- ^ Fish, F.E.; Hui, C.A. (1991). "Dolphin swimming–a review". Mammal Review. 21 (4): 181–195. doi:10.1111/j.1365-2907.1991.tb00292.x.
- ^ Campbell, Neil A.; Reece, Jane B. (2005). Biology, 7th Edition. San Francisco: Pearson - Benjamin Cummings. m/s. 522–523. ISBN 978-0-8053-7171-0.
- ^ Hedenstrom, A.; Moller, A.P. (1992). "Morphological adaptations to song flight in passerine birds: a comparative study". Proceedings of the Royal Society of London B: Biological Sciences. 247 (1320): 183–187. Bibcode:1992RSPSB.247..183H. doi:10.1098/rspb.1992.0026.
- ^ Sacktor, B. (1975). "Biochemical adaptations for flight in the insect". Biochemical Society Symposium. 41 (41): 111–131. PMID 788715.
- ^ Salleh, A. (November 7, 2014). "Insects evolved flight as plants grew taller". ABC. Dicapai pada October 14, 2016.
- ^ Barett, Paul M.; Butler, Richard J.; Edwards, Nicholas P.; Milner, Andrew R. (September 26, 2007). "Pterosaur distribution in time and space: an atlas" (PDF). Zitteliana. B28: 61–107. ISSN 1612-4138.
- ^ Pascal Godefroit; Andrea Cau; Hu Dong-Yu; François Escuillié; Wu Wenhao; Gareth Dyke (2013). "A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds". Nature. 498 (7454): 359–62. Bibcode:2013Natur.498..359G. doi:10.1038/nature12168. PMID 23719374.
- ^ "Vertebrate flight: Chiropteran flight". Dicapai pada October 14, 2016.
- ^ "Merriam's Kangaroo Rat Dipodomys merriami". U. S. Bureau of Land Management web site. Bureau of Land Management. Dicapai pada 2014-03-26.
- ^ Merlin, P. (2014). "Heteromyidae: Kangaroo Rats & Pocket Mice". Arizona-Sonora Desert Museum web site. Arizona-Sonora Desert Museum. Dicapai pada 2014-03-26.
- ^ a b "Animal Guide: Giant Kangaroo Rat". Nature on PBS web site. Public Broadcasting System. 2014. Dicapai pada 2014-03-26.
- ^ Schroder, G.D. (August 1979). "Foraging behavior and home range utilization of the Bannertail Kangaroo Rat". Ecology. 60 (4): 657–665. doi:10.2307/1936601. JSTOR 1936601.
- ^ "Top 10 best jumper animals". Scienceray. Diarkibkan daripada yang asal pada 2009-09-07. Dicapai pada 2012-06-11.
- ^ James, R. S.; Wilson, R. S. (2008). "Explosive jumping: extreme morphological and physiological specializations of Australian rocket frogs (Litoria nasuta)" (PDF). Physiological and Biochemical Zoology. 81 (2): 176–185. doi:10.1086/525290. PMID 18190283.
- ^ Quillan, K.J. (2000). "Ontogenetic scaling of burrowing forces in the earthworm Lumbricus terrestris". Journal of Experimental Biology. 203 (Pt 18): 2757–2770. doi:10.1242/jeb.203.18.2757. PMID 10952876.
- ^ "Underwater bipedal locomotion by octopuses in disguise". Science. 307 (5717): 1927. 2005. doi:10.1126/science.1109616. PMID 15790846.
- ^ Naish, Darren (2008-12-03). "B. rex! – Tetrapod Zoology". Scienceblogs.com. Dicapai pada 2014-06-10.
- ^ Parker, W. K. (1891). "On the morphology of a reptilian bird, Opisthocomus hoazin". Transactions of the Zoological Society of London. 13 (2): 43–89. doi:10.1111/j.1096-3642.1891.tb00045.x.
- ^ "Butterflies in the Nymphalidae family". Dicapai pada 4 October 2016.
- ^ a b Sensenig, Andrew T; Jeffrey W Shultz (2003-02-15). "Mechanics of Cuticular Elastic Energy Storage in Leg Joints Lacking Extensor Muscles in Arachnids". Journal of Experimental Biology. 206 (4): 771–784. doi:10.1242/jeb.00182. ISSN 1477-9145. PMID 12517993.
- ^ Shultz, Jeffrey W (2005-02-06). "Evolution of locomotion in arachnida: The hydraulic pressure pump of the giant whipscorpion, Mastigoproctus Giganteus (Uropygi)". Journal of Morphology. 210 (1): 13–31. doi:10.1002/jmor.1052100103. ISSN 1097-4687. PMID 29865543.
- ^ Shultz, Jeffrey W (1992-01-01). "Muscle Firing Patterns in Two Arachnids Using Different Methods of Propulsive Leg Extension". Journal of Experimental Biology. 162 (1): 313–329. doi:10.1242/jeb.162.1.313. ISSN 1477-9145. Dicapai pada 2012-05-19.
- ^ Quillin KJ (May 1998). "Ontogenetic scaling of hydrostatic skeletons: geometric, static stress and dynamic stress scaling of the earthworm lumbricus terrestris". The Journal of Experimental Biology. 201 (12): 1871–83. doi:10.1242/jeb.201.12.1871. PMID 9600869.
- ^ Cartmill, M. (1985). "Climbing". Dalam M. Hildebrand; D.M. Bramble; K.F. Liem; D.B. Wake (penyunting). Functional Vertebrate Morphology. Belknap Press, Cambridge. m/s. 73–88.
- ^ Fröbisch J.; Reisz, R.R. (2009). "The Late Permian herbivore Suminia and the early evolution of arboreality in terrestrial vertebrate ecosystems". Proceedings of the Royal Society B: Biological Sciences. 276 (1673): 3611–3618. doi:10.1098/rspb.2009.0911. PMC 2817304. PMID 19640883.
- ^ "Portuguese Man-of-War". National Geographic Society. Dicapai pada December 16, 2014.
- ^ "Sharksucker fish's strange disc explained". Natural History Museum. 28 January 2013. Diarkibkan daripada yang asal pada 1 February 2013. Dicapai pada 5 February 2013.
- ^ University of Utah (2008). Ecology and Evolution of Transmission in Feather-feeding Lice (Phthiraptera: Ischnocera). m/s. 83–87. ISBN 978-0-549-46429-7.